Surgery, Gastroenterology and Oncology

|
|
Background: One of the most prevalent solid tumors in men is prostate cancer which is caused by a lack of apoptosis, which leaves cancerous cells that refuse to die. Consequently, triggering apoptosis is a common approach for anticancer treatments to destroy tumor cells. The bacterial biofilm's EPS (Exopolysaccharides) is paying attention as a potential novel cancer treatment source.
Methods: This study targeted to determine the anti-cancer capabilities of EPS extract and its impact on the expression of the genes Bcl-2, Bax, p53, caspase3, and caspase9 in the prostate cancer cell line (LNCaP).
Results: Treatment of LNCaP cells with EPS extract caused a significant increase in the expression rate of the apoptosis genes and behavior LNCaP cells were driven toward cell death.
Conclusion: According to the obtained result the bacterial EPS shows promising results to be considered as an anticancer drug for prostate tumors.
Introduction
Prostate cancer (PCa) is the second most common solid tumor in men and the fifth cause of cancer mortality (1). Hormone-ablation therapy often has an initial positive effect on prostate cancer patients, but later, the disease becomes androgen-independent and resistant to further treatment (2). Although local surgery, chemotherapy, RT, endocrine therapy, and the use of monoclonal antibodies have improved the prognosis, there are still large numbers of patients at risk of relapse and death (3). Bacterium-mediated cancer therapy (BMCT) has attracted much attention because of its considerable benefits (4). Bacteria has been studied as a potential cancer treatment for over a century (5). William B. Coley, a bone surgeon, first reported that injection of a preparation of heat-killed Streptococcus pyogenes could induce tumor regression in patients with bone and soft tissue sarcoma could experience tumor regression by injecting a preparation of heat-killed Streptococcus pyogenes (6,7). These days, BMCT is receiving greater attention because of advancements in medical technology (8). Various bacteria species, including Streptococcus, Bifidobacterium, Clostridium, and Salmonella species have been studied (5). Bacterial biofilms are complex surface attached communities of bacteria held together by self produced polymer matrixes mainly composed of polysaccharides, secreted proteins, and extracellular DNAs (9). To prevent desiccation and maintain adhesions by building biofilms, bacteria release EPS into an environment in the capsules form or slime (10) Water soluble Ionic or nonionic polymers can make up polysaccharides (10). Several factors and para meters influence the production of EPS among these are the composition of the medium, especially carbon and nitrogen sources, pH, temperature, and incubation time (10). EPS exhibit effectiveness as an antioxidant, anticancer, and immune stimulating activities attribute have lead to increase attention to EPS, which is regard as one of a top 10 global reasons of death as substances secrete from S. aureus biofilm might impair immune responses (11). Roca et al. (10) were considered the first who informed the production of EPS with high uronic acids. the EPS has intriguing qualities that make it suitable for applica tion in tissue engineering and regenerative medicine, much like heparin, due to its high uronic acid content. Priyanka et al. (10) showed that the EPS (650 mg l 1) with 7.08% uronic acid containing sugars and sulphate functional group (2.68%); its molecular weight was 90 kDa produced from isolated Nitratireductor sp. strain PRIM 31 is a promising drug for brain tumors through this anionic charged of EPS (the Presence of uronic acid, sulphate functional groups, and phosphate to this EPS assigns an overall anionic charge to the polymer) binding to the epidermal growth factor (EGF) secreted by the tumor and this coincides with the findings of Liu et al. (10) that anionically charge EPS be preventing EGF receptors phosphorylation. The EPS has a sulfur group, which is a crucial mineral needed in precise amounts for healthy metabolism and antioxidant defense mechanisms. Carcinogenic trans formation prevents cysteine from being bio-degraded into sulfane sulfur metabolite, which leads to a disruption in cellular redox control. The triggering of apoptosis and the suppression of cell proliferation depend on the control of cellular redox (10).
Identifying and targeting apoptosis genes has been a major effort in developing effective therapeutic strategies for advanced cancer (8). In this regard mutations in the p53 gene is a common molecular abnormality in cancer (12). A consequence of the lack of normal function of p53 may be the failure to induce apoptosis in cells with damaged DNA, and it can also possibly impair a full apoptotic response to the administration of hormonal or chemotherapeutic interventions (13). Furthermore, the genes of the Bcl 2 family have emerged as key regulators of apoptosis. Several members of the Bcl 2 family, including bcl 2 suppress apoptosis; whereas others, including Bax, induce apoptosis (14). Decreasing anti apoptotic Bcl 2 levels with increasing tumor grade, it has been proposed that Bcl 2 has an early role within the tumor by rescuing cells with otherwise lethal mutations. After additional oncogene activation, some cells would acquire additional ways to protect themselves against apoptosis (15). At that point, loss of Bcl 2 might confer a growth advantage. In fact, Bcl 2 is known to restrain cell proliferation (16)Thus, expression of Bcl 2 would change from high levels in early or low grade tumors, characterized by low apoptotic indices, to low levels in advanced or high grade tumors, characterized by high apoptotic indices (17).
In addition, during apoptotic cell death caspases genes also, play an essential role. The control of caspases is a key and central component of the biochemical pathway, At least three distinct pathways for caspase activation exist in mammalian cells; recruitment activation, trans activation, and auto activation. Trigger caspase activation for the treatment of disorders where insufficient apoptosis occurs (e.g. cancer), is one of the therapeutic strategies (18,19).
This study tested the cytotoxic effects of the EPS from S. aureus against Vero and LNCaP cells, then analyzed the apoptosis related genes in prostate cancers after treatment cells with extract EPS.
Materials And Methods
Bacterial Strain and Culture Medium
S. aureus strains were grown in Tryptic-Soy-Broth (TSB) prepared as per the manufacturer's instructions and autoclaved (121 °C / 15 min).
Quantitative Biofilm Formation
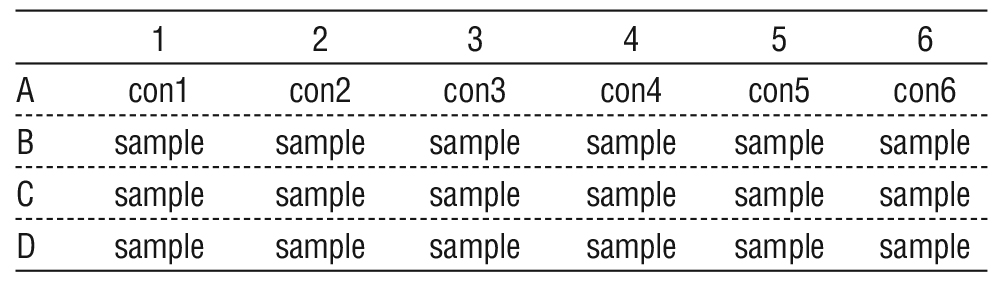
The ability of S. aureus to create biofilm was assessed through adherence to polystyrene microtiter plates. Briefly, 100 ?l of TSB plus supplements were placed into each of the 96 wells of flat bottom sterile polystyrene tissue culture plates. Each well received 100 ?l of a diluted overnight bacterial culture (1:100 in TSB). In negative control wells, only TSB was present. Following a 24 hour incubation period, the plate contents were poured off, and the wells underwent three rounds of deionized water washing. After allowing the microtiter plates to air dry for half an hour, 0.1% w/v crystal violet stain was applied. After properly cleaning the plates with deionized water to remove any remaining stains, they were left to dry at room temperature for two hours. Biofilm generation was quantified by measuring the absorbance at a wave length of 570 nm (OD 570 nm) using a micro ELISA auto reader (20), table 1.
Table 1 - Layout of sample uninoculated medium as (control), and the samples of inoculated medium with S. aureus in micro ELISA.
Congo Red Agar Method
The Congo red agar method is another method of screening biofilm formation, which requires the use of a specially prepared solid medium brain heart infusion broth (BHI) supplemented with 5% sucrose and Congo red (21).
Extraction Crude Exopolysaccharides (EPS)
Following the formation of mature biofilms, the culture broth was centrifuged for 15 minutes at 4,500 g and 4°C, precipitating and discarding the cell pellets. The supernatant at night left in 4°C with 5% Trichloroacetic acid added then all cells were eliminated and no protein content remain. Then centrifuged at 3,500 g, using 0.1 M NaOH, and the pH of the clear supernatant was brought to 7.0. After adding 95% ethanol to each supernatant in three volumes and let it sit at 4 °C for at night, the precipitate of EPS was separated by centrifugation at 3,500 g for 20 min at 4°C. It was then twice cleaned with acetone and dehydrated with ether. Finally, dried in a desiccator then weighed and stored at room temperature (22).
Structural Characterization of the EPS
The dried EPS samples were analyzed by FTIR spectroscopy in ATR mode using an FTIR spectrometer. The spectrum was recorded in transmittance mode in the range of 400 – 4,000 cm1 (23).
Table 2 - Forward and reverse primer sequences

Preparation of EPS Extract as Treatment
To prepare different concentrations of bacterial Exopolysaccharides (EPS) extract of S. aureus was dissolved in sterile RPMI. The obtained solution was filtered twice with the filter paper. Then the solution was sterilized by 0.22 micron filters and concentrations of 20,80, 160, 320, and 640 ?g/ ml) were prepared by serially diluting of standard stock solution (640 ?g/ ml).
Cell Culture and Treatment
Vero cells and human prostate carcinoma cell lines LNCaP cells were purchased from the Pasteur Institute of Iran and maintained in the proper conditions. The Vero cell was maintained in RPMI1640 90%, supplemented with fetal bovine serum (10%), and 1% (100 ?g /ml Penicillin and 100 ?g/ml streptomycin) medium. The LNCaP cells were cultured in 50% Dulbecco’s modified Eagle’s Medium (DMEM) and 50% RPMI 1640 supplemented with 10% fetal bovine serum (FBS), 100 ?g/mL penicillin, and 100 ?g/mL streptomycin (Invitrogen, Carlsbad, CA), at 37 °C and maintained humidified incubator with 5 % CO2. The cells were harvested after trypsinization (0.025 % trypsin and 0.02 % EDTA) and washed twice with phosphate buffered saline (PBS). When the cell density reached approximately 80%, cells were split for further culture. The experiments were made up in the growth phase of cells in the logarithmic (24).
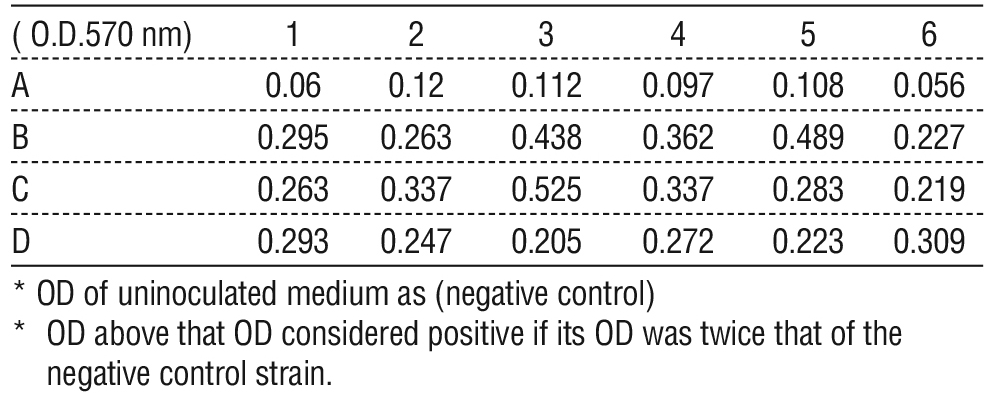
Table 3 - The OD of control samples (A), and (B,C,D) show the OD of S. aurous in micro ELISA.
Cytotoxicity Assay
The EPS effect on proliferative capacity (Viability) was determined using 3 (4,5 dimethylthiazol 2 yl) 2,5 diphenyltetrazolium bromide assay. Briefly, 1 × 104 cells per well (LNCAP, and Vero) were plated in 96 well culture plates. After overnight incubation, the cells were treated with different concentrations of EPS (20, 80, 160, 320 and 640 ?g/ml) for 24, 48, and 72 hours with three replicates. In this assay, the resulting formazan crystals that had formed dissolved in DMSO (150 ?L). Absorbance was recorded at 540 nm with a reference at 650 nm serving as the blank. The EPS effect on cells viability was assessed as % cell viability compared with control on treatment cells, which were arbitrarily assigned 100% viability. This experiment was performed in triplicate and the data were obtained as mean±SD (24,25).
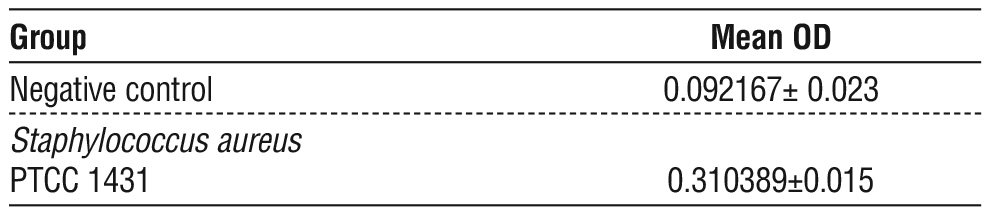
Table 4 - The mean of OD for Staphylococcus aureus and Negative control
Primers Preparation
All primers used in this study were provided by Macrogen in a lyophilized form. as the manufacturer's instructions all lyophilized primers were in 300 µL of nuclease free water to obtain a final concentration of 100 pmol/µL as a stock solution. Working solution was prepared by adding 10 µL of primer stock solution frozen at 20 °C to 90 µL of nuclease free water to give a final concentration of 10 pmol/µL of working primer stock solution.
Primers
The sequences (forward and reverse primer) for RT PCR and Real time PCR were selected according to previously published articles. The primers with reference are shown in table 2.
Figure 1 - Screening of biofilm producers by TCP method: high and non-slime producers differentiated with crystal violet staining in 96 well tissue culture plate.

RNA Extraction and cDNA Synthesis
RNA extraction and cDNA synthesis were performed (30). In brief, the steps would be as follows: (1) extraction of total RNA by SinaPureTM RNA kit from Sinaclon company (2) quantization of RNA’s concentration and purity by NanoDrop 1000 Spectrophotometer (NanoDrop Technologies, Inc. Wilmington, DE, USA), (3) comple mentary DNA (cDNA) synthesis by reverse transcription reaction, by SinaClon First Strand cDNA synthesis kit from Sinaclon company (4) quantization of cDNA concentration by NanoDrop 1000 Spectrophotometer.
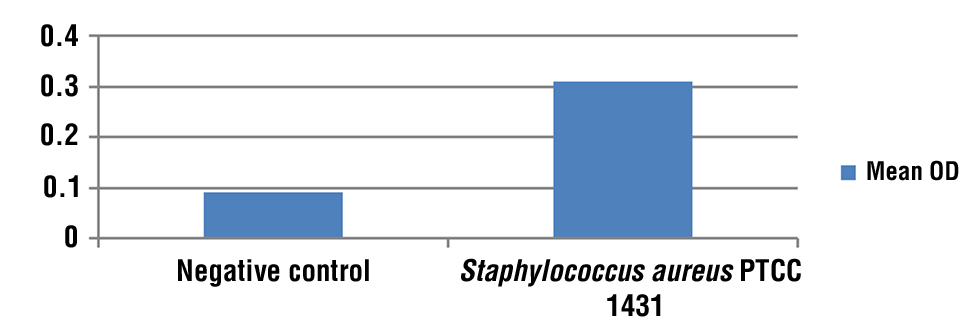
Figure 2 - Graph mean of OD for Staphylococcus aureus and Negative control.
Real-time PCR
Real time PCR was carried out. GAPDH were used as housekeeping genes and were amplified by PCR using forward and reverse primers, table 2 contains a list of all primer genes. Each primer and cDNA template was used with 2X SYBR Green dye, and qPCR Master Mix to create the PCR reactions. The 40 cycles of the PCR reaction were held for 30 seconds at each temperature level after the first cycle of initial denaturation at 94°C for 10 min was completed in the real time PCR program. Finally, melting curve analysis was performed over a gradient extending from an annealing to a denaturation temperature. The expression was calculated by using the relative standard curve method of quantification and reported as a fold change in gene expression. Finally, melting curve analysis was performed over a gradient extending from an annealing to a denaturation temperature. The expression was calculated according to (31).
Figure 3 - Biofilm of S. aureus result on congo red agar plate.

Statistical Analysis
The data were expressed as mean values ± SD. The difference between the control and treated groups was analyzed by One way ANOVA. P<0.05 was considered statistically significant.
Result
Tissue Culture Plate (TCP) Method or (Microtiter Plate Test)
TCP showed positive findings, with a high significant differences (p?0.05). The average OD of S. aureus as (Positive Control B, C, D) was 0.310389 ± 0.015 and the average OD of the uninoculated medium as (negative control A) were 0.092167 ± 0.023 (tables 3, 4), (figs. 1, 2).
Figure 4 - Heavy growth of biofilm after culture bacteria four days.

Congo Red Agar (CRA) Method
The CRA plate was inoculated with the bacteria (S. aureus) overnight culture plate and incubated at 37°C for 24-48 h. A positive result was indicated by black colonies with a dry crystalline consistency While the negative result did not change to black. The experiment was performed in triplicate (fig. 3).
Figure 5 - The EPS after Extraction steps from biofilm.
Exopolysaccharides (EPS) Description
S. aureus showed a great growth on the TSB agar medium after 4 days of incubation at 37°C. and showed opaque, off white with mucoid texture colonies (S. aureus gram positive, non motile forms a biofilm on the bottom of the flask), fig. 4. The crude EPS was obtained from bacteria S. aureus grown under optimum conditions, in the stationary phase of growth, and extracted and purified. The amounts of the EPS produced by bacteria are shown in the fig. 5.
Characterization of EPS Extract
The FTIR spectrophotometer of the EPS table 5, showed many peaks to major functional parts of the EPS, specifically carbonyl compounds. The bands at 4000 3500 cm 1 were referred to as the stretching vibration of O H residue in the EPS. The bands at 2800 3000 cm 1 were a result of the stretching vibration of the Lipid (Ch,CH2,CH3). The bands at 1500 1800 cm 1 were a result of the stretching vibration of the carboxyl group, and C=O. The bands 1500 1800 cm 1 indicated the Proteins from C=O, N H, C N (amideI, amideII) residue in an EPS, sulfate active group SO4 is represent at 1800 1000 cm 1 and 900 125 cm 1 indicate Polysaccharides, nucleic acids C O,C O C,N H (amide III) residue in EPS. The major functional groups of the EPS were identified using an FTIR spectrophotometer. The EPS of S. aureus found to contain a sulfur functional group.
Table 5 - Assignments of principal infrared vibrational signals of the (4000-500 cm-1 ) region of the ART/FT-IR spectrum of the S. aureus biofilm.
Cell Viability
The cell viabilities and survival rates of both cells were significantly declined in a dose dependent manner when they were treated with different concentrations of EPS extract, and significantly increased numbers of apoptotic cells.
Cell Viability of Impact of S. aureus EPS on Vero Cell Line
Results showed there were significant differences in the Vero cell lines viability, dose dependent manner was observe in cells treat with a EPS of biofilm. The greater rates of Vero cell viability was at a concentration of 20 µg/ml and the lowest rate of viability was at a concentration of 640 µg/ml at different time treatment exposure (24-, 48-, 72- hrs), and there was remarkable inhibitions in cells viability represente by a marked inhibitory effects on the cell population fig. 6, also there were no significant differences between viability time dependent manners, table 6.
Figure 6 - Steady decline of the Vero cell population that noted after treated with IC50 dose of EPS at 48h as compared to the control (Untreated cells).

Cell Viability of Impact of EPS on LNCaP Cell Line
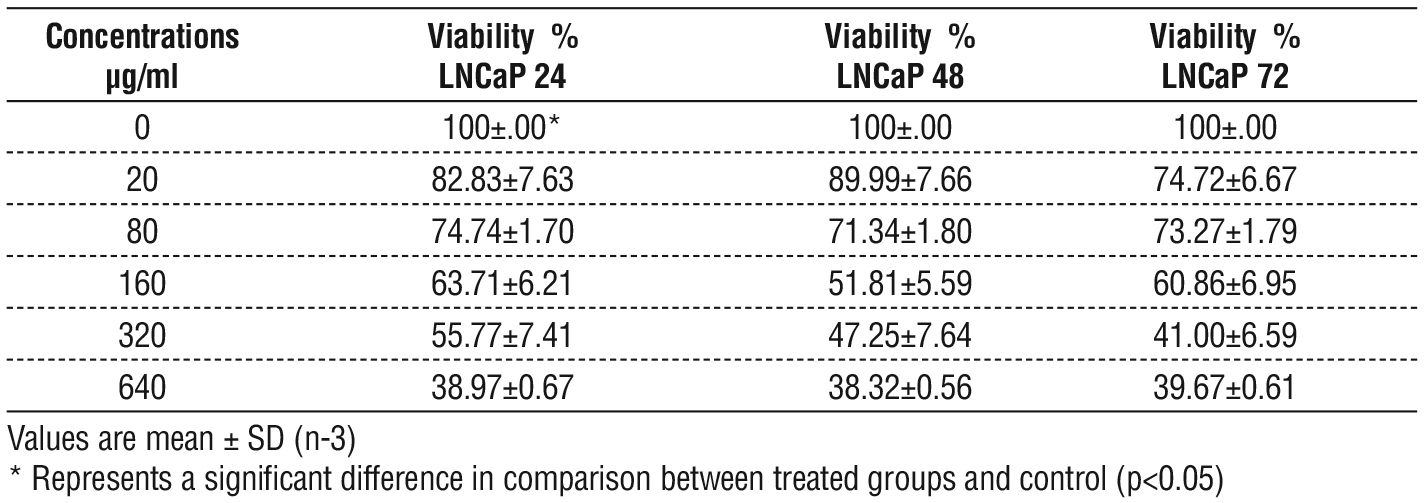
Results showed significant differences in the viability of the LNCaP cell lines, dose dependent manner were observe in cells treat with EPS. The greater rate of viability LNCaP cell was at a concentra tion of 20 µg / ml and the fewer rate of viability was at a concentration of 640 µg/ml. At different times of treatment exposure (24, 48, 72 hrs) and there was remarkable inhibition in cell viability represented by a marked inhibitory effect on the cell population fig. 7 and table 7. Also, there were no significant differences between the treatments base on time 48-h and 72-h, but there was a clear difference between them and time 24.
Table 6 - Vero cells viability as determined by MTT, post treatment with different concentration of EPS for three times exposure.
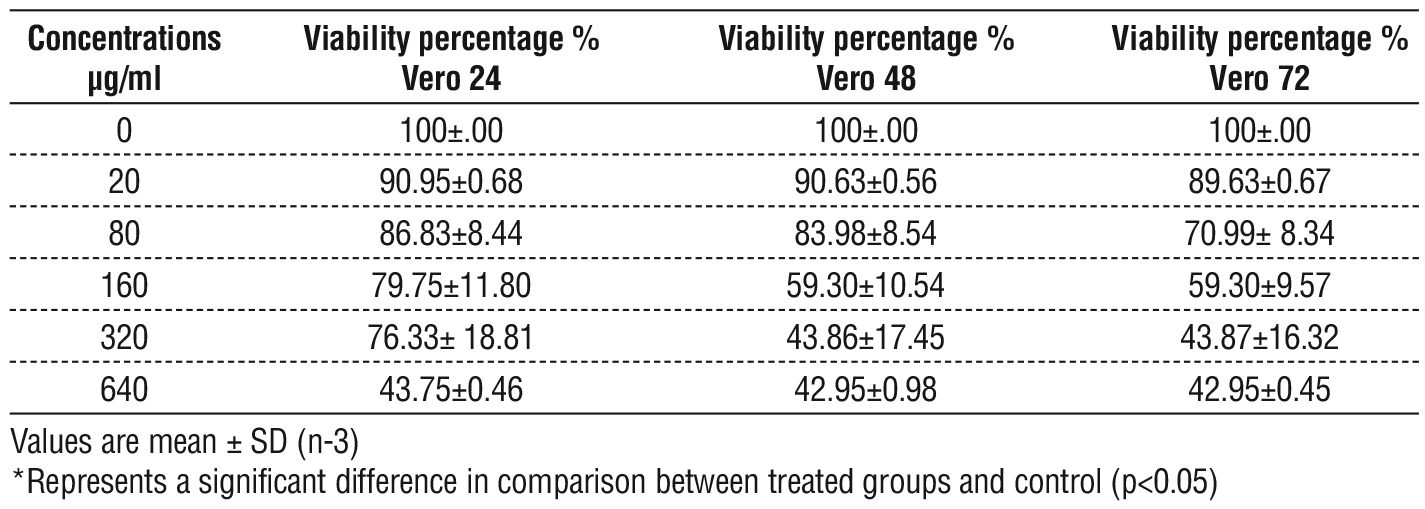
Figure 7 - Steady decline of the LNCaP cell population that noted after treated with IC50 dose of EPS at 48h as compared to the control (Untreated cells)

Table 7 - Viability percentage of LNCaP cell lines
Assess the Half Maximal Inhibitory Concentration (IC50 ) of EPS
The IC50 values were calculated by using the MS Excel curve represented in table 8 and the most active dose of EPS that inhibit growths and proliferation of the VERO and LNCaP cell lines were on 24-h. The IC50 value of Vero cell lines were 479.0123 µg/ml, 454.7238µg/ml and 428.9419µg/ml respectively. While IC50 of LNCaP cell lines value were 442.3609 µg/ml, 252.8518 µg/ml and 206.4241 µg/ml respectively. Also, there was significant difference in IC50 between the time of exposure.
Table 8 - Comparison of IC50 of Vero and LNCaP Cell Lines at three time.

Biochemical Analysis of Cell Death by Real Time PCR
The effect of the tested bacterial EPS on gene expression of cell lines was analyzed by qPCR in order to evaluate the genes expression (P53, BAX, BCL 2, Caspase 9, Caspase 3) in Vero and LNCaP cells was treated with IC50 does of EPS for 48 h (table 9), the RNA samples that were extracted from cells treated with EPS and were reverse transcribed to make the first strand of DNA were used as the templates. cDNA template was mixed with primers amplifying the gene to measure its expression using qPCR relative expression assay using SYBR green master mix. Data were analyzed by ?? CTs and normalized to (GAPDH) house keeping gene were (P < 0.05). The experiment was performed in triplicate.
Table 9 - Gene expression both Vero and LNCaP after treatment with EPS

Figure 8 - The relative expression of P53 in Vero and LNCaP cell treated and untreated with EPS compare to control group.

Apoptosis Induction by Activation p53 Gene
The result found that EPS treatment enhanced the expression of P53 in both cell lines compared to its level in the corresponding controls. Expression was found to have the highest effect of EPS on treated LNCaP cell lines compared to VERO cell lines, fig. 8.
Figure 9 - The relative expression of BAX in Vero and LNCaP cell treated and untreated with EPS compare to control group.

Apoptosis Induction by Activation BAX Gene
The result found that EPS treatment enhanced the expression of BAX in both cell lines compared to its level in the corresponding controls. Expression was found to have the highest effect of EPS on the treated VERO and LNCaP, fig. 9.
Apoptosis Induction by Activation Bcl-2 Gene
The finding found that the expression of Bcl 2 gene showed a significant decrease (P < 0.05) in cancer cell lines (LNCaP) compared to its level in the corresponding controls, On the other hand, treated Vero cell lines didn’t show any significant decrease in bcl 2 gene level, fig. 10.
Figure 10 - The relative expression of Bcl2 in Vero and LNCaP cell treated and untreated with EPS compare to control group.

Ratio of BAX and Bcl2 Genes
TThe obtained data showed that the ratio of Bax/Bcl 2 was increased significantly in LNCaP cells treated with IC50 at 48 hours, compared to the control group and the columnar chart for changes in the relative expression of Bax and Bcl 2 genes in both groups untreated (control) and treated, on the other hand in Vero cell line didn’t showed any significant difference between treated and untreated (control) group, fig. 11.
Figure 11 - The relative expression of BAX and Bcl2 ratio in Vero and LNCaP cell treated and untreated with EPS compare to control group.

Apoptosis Induction by Activation Caspase-3 Gene
The results found that the effect of the prepared EPS on the expression of Caspase 3 gene was signifi cantly increased in treated cancer cells LNCaP with at IC50 does at 48 h, compared to the control sample, while the VERO showed a slight increase in the gene, fig. 12.
Figure 12 - The relative expression of Caspase-3 in Vero and MCF-7 cell treated and untreated with EPS compare to control group.

Apoptosis Induction by Activation Caspase-9 Gene
The results of Caspase 9 expression showed that the gene was higher in normal cells (Vero) compared to the negative control sample and the cancer (LNCaP), the LNCaP showed a slight increase in the gene was observed compared to the negative control sample, fig. 13.
Figure 13 - The relative expression of Caspase-9 in Vero and LNCaP cell treated and untreated with EPS compared to control group.

Discussion
Bacterial biofilms are complex surface attached bacterial populations kept together by self produced polymer matrixes primarily consisting of Exopoly saccharides EPS, EPS accounts for around 50% to 90% of the total organic carbon in biofilms and can be regarded as the major matrix material of the biofilm. Although the chemical and physical properties of EPS vary, it is mostly made of polysaccharides. The (EPS) plays a critical role in bacterial, fungal, and algal defense systems (32). In recent years, EPSs have had great potential as anti tumor drugs as recent treatment strategies, so the study considers it of interest to extract EPS from bacteria and used it as anti tumor effects on prostate cancer carcinoma.
In current study's findings demonstrated that cells treated with EPS exhibited suppression of cell viability in a concentration dependent manner and LNCaP was shown to be the most sensitive cell line to EPS, whereas VERO was the most tolerant.
Authors reported that EPS produced by bacteria strains has cytotoxic action on cancer cells by inhibiting growth and proliferation of the HepG2 cell line, these crude EPSs were obtained from bacteria grown under optimum conditions, in the stationary phase of growth, and extracted and purified (22). Others observed nearly the same result, they found that E. lactis IW5 secretions had no harmful effects on healthy cells (33).
Priya JN et al (34) showed that the EPS with 7.08% uronic acid containing sugars and sulfate functional group, is a promising drug for brain tumors through this anionic charged EPS (the Presence of uronic acid, sulfate functional groups, and phosphate to this EPS assigns an overall anionic charge to the polymer) binding to the epidermal growth factor (EGF) secreted by the tumor and this coincides with the findings of (33), that EPS with an anionic charge has the ability to stop the phosphorylation of the EGF receptor, with EPS (300µg ml) produced cytotoxicity against U87MG glioplastoma cells using the MTT assay, and the IC50 value (234.04 µg ml) was demonstrated.
Based on the current characteristics of biofilm and its anti tumor effect, exopolysaccharides (EPS) are crucial components of defense systems in bacteria (34). Vidhyalakshmi and Vallinachiyar (35) also extracted exopolysaccharide from Bacillus that showed cytotoxicity against MCF 7 breast cancer cells without any cytotoxicity against normal cells.
Authors evaluated the anti cancer impact of L. acidophilus was performed with verification of the likely effect of its polysaccharides extract on the suppression of colon cancer cell proliferation (87.27%) and cell death (80.65%) with 1.96–51.3 colon cancer selectivity index (36). The utilized pentasaccharide demonstrated a strong cancer cell selectivity index, reaching 51.3, suggesting that the inhibitory effects of L. acidophilus LA EPS 20079 on colon cancer cell proliferation could be accompanied by an increase in the proliferation of non cancerous cells.
The creation of treatments that encourage the efficient death of cancer cells by apoptosis has been a cornerstone and objective of clinical oncology for more than thirty years. There are two routes that mediate this process of programmed cell death (called intrinsic and extrinsic). Consequently, apoptosis is a mechanism that several anticancer medications used to killing tumor cells, make it a valuable targets to target (37).
The majority of human cancers, if not all of them, include TP53 (p53) gene dysfunction. Due to its nearly ubiquitous inactivation in cancer, p53 is a very desirable target for the development of novel anti cancer medications. P53 can induce apoptosis through both extrinsic and intrinsic mechanisms (38) and many cancer treatment induced apoptosis are mediate by the tumor protein p53 activation, Bax and sub sequent activation of caspases (39).
Normal cells that are not under stress have very low levels of p53 and very little of it active. Stress causes p53 to undergo a number of post translational changes that activate it and allow it to attach to particular DNA sequences (13). Numerous hundred genes that are variably regulated (induced or repressed) based on the type of cell include the very loose p53 recognition sequence (13). The kind of stress and the degree of harm, The findings demonstrated that the EPS extract had a significantly distinct effect on P53 expression; the treated LNCaP had the highest expression followed by treated Vero cells. Consequently, our findings suggested that EPS reduced the mortality of prostate cancer cell death by raising p53.
A study found that the mitochondrial pathway was implicated in the EPS induced apoptosis due to the up regulation of Bax and p53 mRNA and the down regulation of Bcl 2 mRNA (40). These results shed new light on the possible application of EPS as an anticancer medication in the treatment of human colo-rectal cancers.
According to Sangour and By (2020), Zns Ag NPs treatment (25 ?g/ml) for 48 hours boosts P53 expression levels in both treated cell lines (VERO and MCF 7) when compared to the control, with treated samples of MCF 7 showing a notable rise. Since treating three different types of cell lines with ZnS impacts their viability and increases the P53 mRNA level, P53 upregulates expression is consistent with Akhtar et al (35). On the other hand, P53 mRNA increases almost 35 times as opposed to twofold. This could be explained by the concomitant action of the utilized nanoparticles' Ag NPs.
P53 expression in MCF 7 treatment is 16 times higher than in control, with relative increases in P53 in VERO cells, after 48 hours of treatment with an IC50 concentration of CdS Au NPS in VERO and MCF 7 cells. According to a different study, reactive oxidizing species and nanoparticles damage DNA and activate P53 and other proteins involved in DNA repair (41). The p53 plays a controlling gene linked to apoptosis In addition to that it also seem to have a direct effect on mitochondria, potentially encourage the convergence of extrinsic and intrinsic pathways. It can interacting with members of the BCL2 family to lead to apoptosis and mitochondrial outer membrane permeabilization (35).
Apoptosis characteristics that have been studied to target cell death therapeutically include the release of cytochrome C from the mitochondria (42), which is controlled by the balance between pro apoptotic BAX and anti apoptotic BCL 2 family members (16). Our data found that the expression of Bax and Bcl 2 genes was also associated with a significant increase in IC50 doses of EPS extract at 48 hours in treated LNCaP cell lines, compared to cells that were not treated with the EPS extract. On the other hand.
The Bax/Bcl2 ratio determines whether a cell lives or dies. Some chemicals can change the Bax/Bcl 2 ratio, causing cancer cells to die (39). A study on anticancer pharmaceuticals discovered that these drugs change the Bax/Bcl 2 ratio in the AGS cell line, causing malignant cells to die. Some plant compounds can induce cancer cells to die by increasing the Bax/Bcl 2 ratio. Pyrethrin is a naturally occurring insecticide that boosts the Bax/Bcl 2 ratio in HepG2 cells (a human liver cancer cell line) (43). in the present study when comparing untreated cells to treat MCF 7 cell lines that expressed the Bax and Bcl 2 genes had significantly more (P< 0.05) IC50 dosages of EPS extract after 48 hours. As a result, cancerous cells commit suicide.
Nowadays the development of anti cancer drugs has been shifted toward cell cycle transduction signals, growth factors, DNA repair, and apoptosis. Apoptosis can occur via two pathways: the extrinsic pathway and the intrinsic pathway (42). When mitochondria are damaged, the intrinsic apoptotic pathway is activated. Caspase 9 acts as a starter in the intrinsic pathway of apoptosis activation, while caspase 3 acts as an "executor" enzyme. As a result, an increase in caspase3 leads to an increase in apoptosis. Furthermore, altered caspase 3 expression may represent an additional route of androgen resistance (44).
The study was conducted in vitro on prostate cancer cell lines (LNCaP), when comparing the treated LNCaP cell line to the control (untreated cells), the current study found a considerable rise in Caspase 3 and a slight increase in Caspase 9. These findings suggest that EPS may have therapeutic potential in preventing the growth of prostate tumors. However, there was also an increase in both Caspase 3 and Caspase 9 in the Vero cell lines that were treat compared to those not treated. Caspase is a mediatory enzyme involved in cellular apoptosis; thus, increased caspase expression is an indicator of an ongoing apoptosis process in cancer cells (45). Numerous proteins/enzymes mediate cell death by the intrinsic pathway (caspase 9) and caspase executor (caspase 3) (46). A study by Zhang et al. (29) demonstrated that caspase 3 was a crucial component in the apoptotic signaling linked with the activation of internucleosomal DNA fragmentation. A study demonstrated higher expression of caspase 3, ?8, and ?9 that was associated with enhanced apoptosis in cancer cells (47).
A study showed that The increase of caspase 3 can activate specific proteins in cells which will further induce the apoptosis process (48). The increase of caspase 3 enzyme expression will increase the turnover of prostate cancer cell line PC 3 in vitro through the intrinsic pathway. The research presented stated that Cytochrome C is released through an intrinsic mechanism that increases the ratio of Bax to Bcl 2 expression (Bax/Bcl 2) (17). As a result, caspase activation proteins, can either be effectors (caspases 3, 6, and 7) or initiators (caspases 8, 9, and 10) of apoptosis. The apoptosome forms when caspase activation occurs. This study presented similar results to those who stated that Initiator caspases trigger effector caspases, which cause apoptosis by cleaving 100s of proteins via proteolysis (19).
Conclusion
S. aureus extracellular polymer (EPS) has highly promising effects on prostate cancer. However, in vivo investigations are necessary to determine potential interactions between the bacterial EPS and the host immune system, ensuring the safety and efficacy of the EPS in the prevention or treatment of prostate cancer. Additional investigation is necessary to clarify.
Conflict of interests
All authors declared no any conflict of interesting.
Funding
None.
Ethics of approval
This work approved by Biology Department, College of Science, University of Basrah (no. 45 in December 2023).
Reference
1. Gandaglia G, Leni R, Bray F, Fleshner N, Freedland SJ, Kibel A, et al. Epidemiology and Prevention of Prostate Cancer. Eur Urol Oncol. 2021;4(6):877-892.
2. Nicolopoulos K, Moshi MR, Stringer D, Ma N, Jenal M, Vreugdenburg T. The clinical effectiveness of denosumab (Prolia®) in patients with hormone-sensitive cancer receiving endocrine therapy, compared to bisphosphonates, selective estrogen receptor modulators (SERM), and placebo: a systematic review and network meta-analysis. Arch Osteoporos. 2023;18(1):18.
3. Wilkins LJ, Tosoian JJ, Sundi D, Ross AE, Grimberg D, Klein EA, et al. Surgical management of high-risk, localized prostate cancer. Nat Rev Urol. 2020;17(12):679-690.
4. Sarotra P, Medhi B. Use of Bacteria in Cancer Therapy. Recent Results Cancer Res. 2016;209:111-121.
5. Badie F, Ghandali M, Tabatabaei SA, Safari M, Khorshidi A, Shayestehpour M, et al. Use of Salmonella Bacteria in Cancer Therapy: Direct, Drug Delivery and Combination Approaches. Front Oncol. 2021;11:624759.
6. Kahrarian Z, Horriat M, Khazayel S. A Review of Novel Methods of the Treatment of Cancer by Bacteria. 2019;1(1):25–36.
7. Mager, D. Bacteria and cancer: cause, coincidence or cure? A review. J Transl Med 2006;4:14.
8. Duong MT, Qin Y, You SH, Min JJ. Bacteria-cancer interactions: bacteria-based cancer therapy. Exp Mol Med. 2019;51(12):1-15.
9. Alboslemy T, Yu B, Rogers T, Kim MH. Staphylococcus aureus Biofilm-Conditioned Medium Impairs Macrophage-Mediated Antibiofilm Immune Response by Upregulating KLF2 Expression. Infect Immun. 2019;87(4):e00643-18.
10. Wijesekara I, Pangestuti R, Kim S-K. Biological activities and potential health benefits of sulfated polysaccharides derived from marine algae. Carbohydr. Polym. 2011;84(1):14–21.
11. Josse J, Valour F, Maali Y, Diot A, Batailler C, Ferry T, et al. Interaction Between Staphylococcal Biofilm and Bone: How Does the Presence of Biofilm Promote Prosthesis Loosening? Front Microbiol. 2019; 10:1602.
12. Williams AB, Schumacher B. p53 in the DNA-Damage-Repair Process. Cold Spring Harb Perspect Med. 2016;6(5):a026070.
13. Rivlin N, Brosh R, Oren M, Rotter V. Mutations in the p53 Tumor Suppressor Gene: Important Milestones at the Various Steps of Tumorigenesis. Genes Cancer. 2011;2(4):466-74.
14. Kunac N, Filipovi? N, Kosti? S, Vukojevi? K. The Expression Pattern of Bcl-2 and Bax in the Tumor and Stromal Cells in Colorectal Carcinoma. Medicina (Kaunas). 2022;58(8):1135.
15. Warren CFA, Wong-Brown MW, Bowden NA. BCL-2 family isoforms in apoptosis and cancer. Cell Death Dis. 2019;10(3):177.
16. Singh R, Letai A, Sarosiek K. Regulation of apoptosis in health and disease: the balancing act of BCL-2 family proteins. Nat Rev Mol Cell Biol. 2019;20(3):175-193.
17. Qian S, Wei Z, Yang W, Huang J, Yang Y, Wang J. The role of BCL-2 family proteins in regulating apoptosis and cancer therapy. Front Oncol. 2022;12:985363.
18. Ameen AA, Suker DK. MIR30a suppresses prostate cancer cells by increasing apoptosis and regulation of MMPs, Eurasian Med. Res. Period. 2022;9:26–34.
19. Nicholson DW. Caspase structure, proteolytic substrates, and function during apoptotic cell death. Cell Death Differ. 1999;6(11): 1028-42.
20. Malik N, Bisht D, Aggarwal J, Rawat A. Phenotypic Detection of Biofilm Formation in Clinical Isolates of Methicillin-Resistant Staphylococcus aureus. Asian J. Pharm. Res. Heal. Care. 2022; 14(1):43.
21. Hassan A, Usman J, Kaleem F, Omair M, Khalid A, Iqbal M. Evaluation of different detection methods of biofilm formation in the clinical isolates. Braz J Infect Dis. 2011;15(4):305-11.
22. Abdelnasser SM, M Yahya SM, Mohamed WF, Asker MM, Abu Shady HM, Mahmoud MG, et al. Antitumor Exopolysaccharides Derived from Novel Marine Bacillus: Isolation, Characterization Aspect and Biological Activity. Asian Pac J Cancer Prev. 2017; 18(7):1847-1854.
23. Rehman ZU, Vrouwenvelder JS, Saikaly PE. Physicochemical Properties of Extracellular Polymeric Substances Produced by Three Bacterial Isolates From Biofouled Reverse Osmosis Membranes. Front Microbiol. 2021;12:668761.
24. Freshney RI. Culture of animal cells: a manual of basic technique and specialized applications. John Wiley & Sons, 2015; p. 123-177.
25. Mazyed ZA, Suker DK, Tawfeeq AT. Antioxidant, Anti-tumor activity of cichorium intybus extracts on two cell lines in vitro. Biochem. Cell. Arch. 2021;21(2):1-6.
26. Bosso M, Al-Mulla F. RKIP & GSK3b: The interaction of two intracellular signaling network regulators and their role in cancer. In: Prognostic and Therapeutic Applications of RKIP in Cancer, Elsevier, 2020, p. 147–173.
27. Xiong C, Li P, Luo Q, Phan CW, Li Q, Jin X, et al. Induction of Apoptosis in HeLa Cells by a Novel Peptide from Fruiting Bodies of Morchella importuna via the Mitochondrial Apoptotic Pathway. Evid Based Complement Alternat Med. 2021;2021:5563367.
28. Yaoxian, W., Hui, Y., Yunyan, Z. et al. Emodin induces apoptosis of human cervical cancer hela cells via intrinsic mitochondrial and extrinsic death receptor pathway. Cancer Cell Int. 2013; 13: 71. https://doi.org/10.1186/1475-2867-13-71.
29. Zhang Y, Ge Y, Shi W, Hu C, Bai X, Rong J, Yu M. CDS-1548 induces apoptosis in HeLa cells by activating caspase 3. Oncol Lett. 2019; 18(2):1881-1887.
30. Mitupatum T, Aree K, Kittisenachai S, Roytrakul S, Puthong S, Kangsadalampai S, et al. Hep88 mAb-mediated paraptosis-like apoptosis in HepG2 cells via downstream upregulation and activation of caspase-3, caspase-8 and caspase-9. Asian Pac J Cancer Prev. 2015;16(5):1771-9.
31. Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods. 2001;25(4):402-8.
32. Singh AK, Prakash P, Achra A, Singh GP, Das A, Singh RK. Standardization and Classification of In vitro Biofilm Formation by Clinical Isolates of Staphylococcus aureus. J Glob Infect Dis. 2017; 9(3):93-101.
33. Nami Y, Haghshenas B, Yari Khosroushahi A. Molecular Identification and Probiotic Potential Characterization of Lactic Acid Bacteria Isolated from Human Vaginal Microbiota. Adv Pharm Bull. 2018;8(4):683-695.
34. Priya JN, M. Kannan M, P. Priyanka P, Mary SV. A study on characterization of EPS and media optimization for bacterial calcium carbonate precipitation. Int J Curr Microbiol Appl Sci, 2016;5:590–595.
35. Vidhyalakshmi R, Vallinachiyar C.Apoptosis of human breast cancer cells (MCF-7) induced by polysacccharides produced by bacteria. J Cancer Sci Ther, 2013;5(2):31–34.
36. El-Deeb NM, Yassin AM, Al-Madboly LA, El-Hawiet A. A novel purified Lactobacillus acidophilus 20079 exopolysaccharide, LA-EPS-20079, molecularly regulates both apoptotic and NF-kB inflammatory pathways in human colon cancer. Microb Cell Fact. 2018;17(1):29.
37. Carneiro BA, El-Deiry WS. Targeting apoptosis in cancer therapy. Nat Rev Clin Oncol. 2020;17(7):395-417.
38. Chen F, Wang W, El-Deiry WS. Current strategies to target p53 in cancer. Biochem. Pharmacol. 2010;80(5):724–730.
39. Korbakis D, Scorilas A. Quantitative expression analysis of the apoptosis-related genes BCL2, BAX and BCL2L12 in gastric adenocarcinoma cells following treatment with the anticancer drugs cisplatin, etoposide and taxol. Tumor Biol. 2012;33(3):865–875.
40. Yu W, Chen G, Zhang P, Chen K. Purification, partial characterization and antitumor effect of an exopolysaccharide from Rhizopus nigricans. Int J Biol Macromol. 2016;82:299-307.
41. Aldughaim MS, Al-Anazi MR, Bohol MFF, Colak D, Alothaid H, Wakil SM, et al. Gene Expression and Transcriptome Profiling of Changes in a Cancer Cell Line Post-Exposure to Cadmium Telluride Quantum Dots: Possible Implications in Oncogenesis. Dose Response. 2021; 19(2):15593258211019880.
42. Boice A, Bouchier-Hayes L. Targeting apoptotic caspases in cancer. Biochim Biophys Acta Mol Cell Res. 2020;1867(6):118688.
43. Kazemi N, Shahrestani SB. Effect of saffron extract on expression of Bax and Bcl-2 genes in gastric adenocarcinoma cell line (AGS), Gene, Cell Tissue, 2018;5(3):1-5.
44. Elmore S. Apoptosis: a review of programmed cell death. Toxicol Pathol. 2007;35(4):495-516.
45. Fiandalo MV, Kyprianou N. Caspase control: protagonists of cancer cell apoptosis. Exp Oncol. 2012;34(3):165-75.
46. Jiao C, Chen W, Tan X, Liang H, Li J, Yun H, et al. Ganoderma lucidum spore oil induces apoptosis of breast cancer cells in vitro and in vivo by activating caspase-3 and caspase-9. J Ethnopharmacol. 2020;247:112256.
47. Yu CX, Zhang XQ, Kang LD, Zhang PJ, Chen WW, Liu WW, et al. Emodin induces apoptosis in human prostate cancer cell LNCaP. Asian J Androl. 2008;10(4):625-34.
48. Ismy J, Sugandi S, Rachmadi D, Hardjowijoto S, Mustafa A. The Effect of Exogenous Superoxide Dismutase (SOD) on Caspase-3 Activation and Apoptosis Induction in Pc-3 Prostate Cancer Cells. Res Rep Urol. 2020;12:503-508.
Full Text Sources:
Abstract:
Views: 1240

For Authors
Journal Subscriptions

Mar 2026
Supplements
Instructions for authors
Online submission
Contact
e-ISSN: 2601 - 1700 (online)
ISSN-L: 2559 - 723X
Journal Abbreviation: Surg. Gastroenterol. Oncol.
Surgery, Gastroenterology and Oncology (SGO) is indexed in:
- SCOPUS
- EBSCO
- DOI/Crossref
- Google Scholar
- SCImago
- Harvard Library
- Open Academic Journals Index (OAJI)
Surgery, Gastroenterology and Oncology (SGO) is an open-access, peer-reviewed online journal published by Celsius Publishing House. The journal allows readers to read, download, copy, distribute, print, search, or link to the full text of its articles.
Time to first editorial decision: 25 days
Rejection rate: 61%
CiteScore: 0.2

Meetings and Courses in 2025
Meetings and Courses in 2024
Meetings and Courses in 2023
Meetings and Courses in 2022
Meetings and Courses in 2021
Meetings and Courses in 2020
Meetings and Courses in 2019
Verona expert meeting 2019

Surgery, Gastroenterology and Oncology applies the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits readers to copy and redistribute the material in any medium or format, remix, adapt, build upon the published works non-commercially, and license the derivative works on different terms, provided the original material is properly cited and the use is non-commercial. Please see: https://creativecommons.org/licenses/by-nc/4.0/
Publisher’s Note:
The opinions, statements, and data contained in article are solely those of the authors and not of Surgery, Gastroenterology and Oncology journal or the editors. Publisher and the editors disclaim responsibility for any damage resulting from any ideas, instructions, methods, or products referred to in the content.